Independent researcher at SPACE (Lund University)
I recently became a PI and started my own research group! We will continue studying how plants interact with their pollinators on different study systems within the framework of this project:
Drivers of selection on flower traits in a spatio-temporal context
The main aim is to identify the most important biotic and abiotic factors that drive selection in floral traits across altitudes and seasons in the tropical Andes of Bolivia.
CONCEPTUALIZATION (A) and HYPOTHESIS (B):

Evolvability a meta-analyses
In this project we aim to test the evolvability of traits in a global way.
This is work in progress, updates soon.
Postdoctoral research at Sapir lab (TAU University)
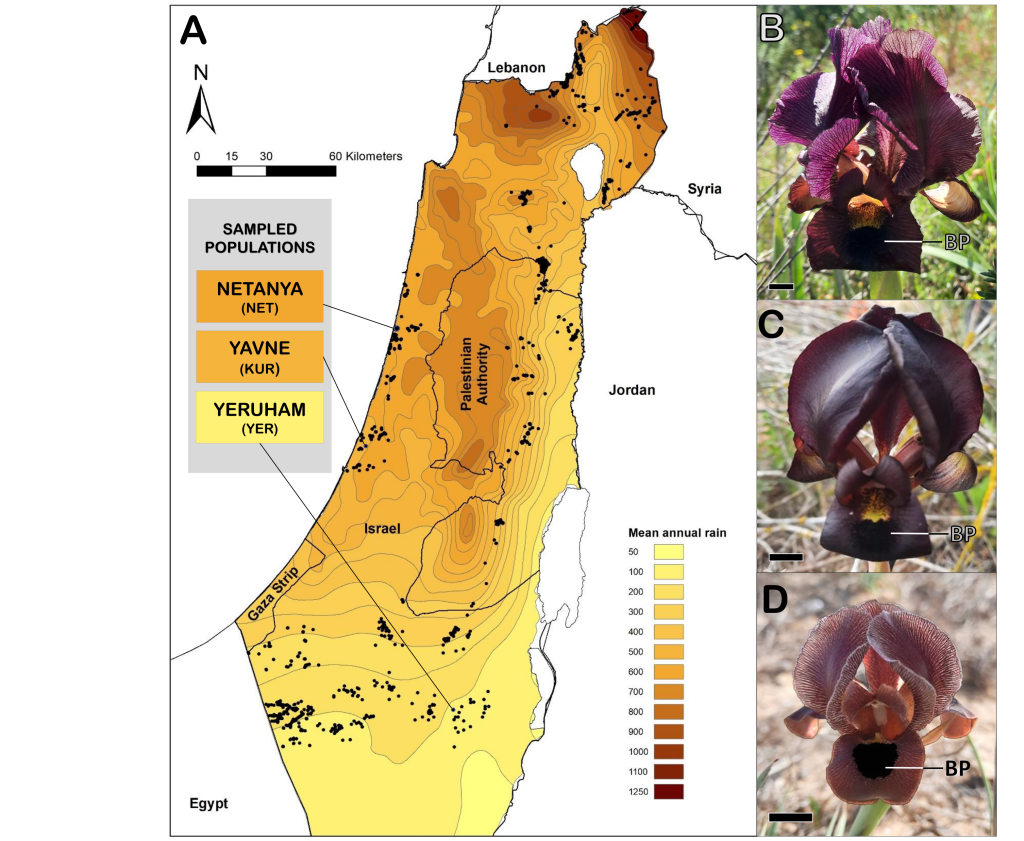
Study system: Royal irises
Royal irises (section Oncocyclus) are endemic from the Middle East. The flowers are very big with three “stand-up” petals called standards and three down petals called falls. In the fall petals they have a characteristic black spot. They are self-incompatible, so they depend on pollinators for their reproduction. However, unlike their close relative, Royal irises do not produce nectar. Eucera long-atenna wild bees pollinate these irises, at sunset the bees look for the flowers to spend the night, the irises offer a shelter to the bees overnight. The next day the flowers heated faster than outside providing heat as a reward to the bees and helping them to fly away faster.






Flower size as a honest signal
Large flowers of Royal irises offer overnight shelter as a reward to Eucera bees. Larger flowers could be easily detected from a distant, while the black patch might signal the entrance to the pollination tunnel (shelter). In this study we tested whether flower size and black patch size could act as a honest signals, indicating larger rewards (larger shelters). We found that flower size and black patch size are positively correlated with tunnel volume (as an extent of the reward) increasing fruit and seed set, suggesting that flower/patch size act as honest signals.
Even though, this positive relationship was found in the entire group of Royal irises (seven species) under controlled conditions, it was only found in YER (Iris petrana) and NET (Iris atropurpurea) in the wild, suggesting that flower/patch size act as honest signals, but its effect on fitness is species or population-specific.
Flower size increased the fitness in YER I. petrana, and interactions between flower/patch size and tunnel size increased fitness in YER and NET populations. This suggests that the honesty of the signal is positively selected in these two populations.




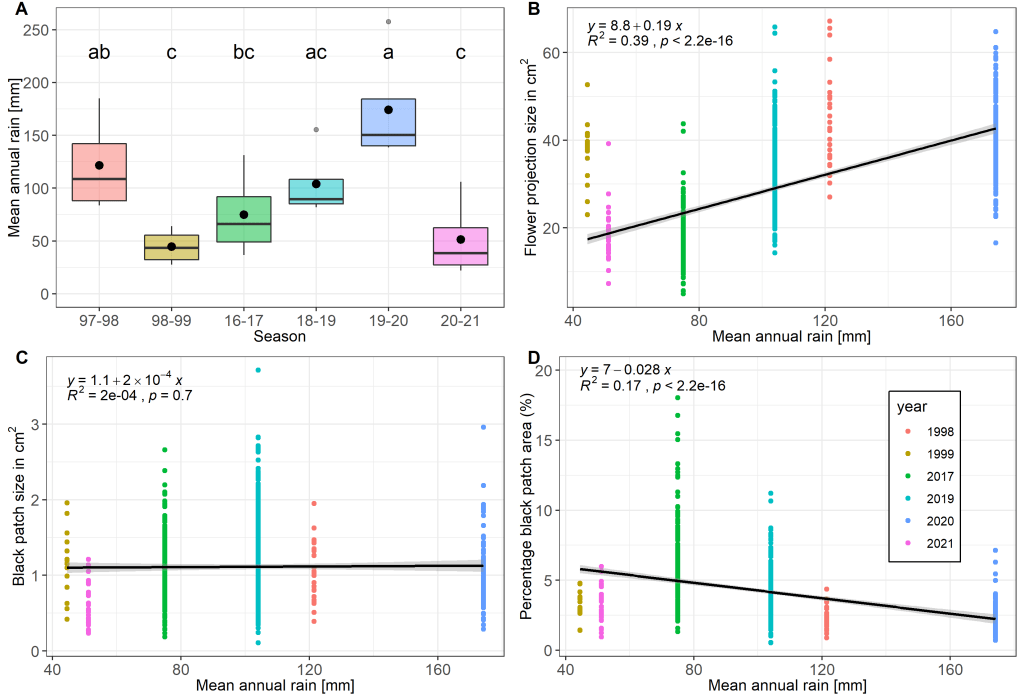
Rainfall variation is related to flower size but not to black patch signal
We studied how the climate (rainfall) affect flower size and black patch size over time in Iris petrana. We found that flower size is highly correlated with rainfall over the years, while black patch size was constant.
These results suggest that the plant is investing resources to keep the area of the black patch constant. This might be an indication that flower size and the black patch size are under different selection regimes. Although flower size might be under a putatively balancing selection of both pollinators and climate, the black patch might be putatively selected by pollinators only.
Flower size and heat reward in Royal irises
Trade-off between flower advertising and flower heat accumulation in Royal irises
We studied the relation between flower size and heat reward in Royal irises (Iris atropurpurea and Iris petrana). We extensively measured flower size in three populations (NET, KUR, YER) and we found a decrease in size from north to south which correlates with the decrease in annual precipitation.
In addition, we measured the temperature inside the flower before and after sunset and tested whether there is a positive correlation with the flower size. The hypothesis was that the size of the flower increases providing for a bigger shelter, and if so also the heat reward should increase.
In contrast to our expectation, the temperature of the flowers increased more rapidly in smaller flowers. We further tested whether the characteristic black patch plays a role in the heating of the flower but we did not find results to support this hypothesis.

This work is currently being consider (major revision) for publication at American Journal of Botany.
Phylogeography of Royal irises
I am also part on this project about the evolutionary history of Royal irises. The main aim is to identify which mechanisms affected the speciation of this group. Using RADSeq we reconstructed the phylogeny based on SNPs, performed IBD (Isolation by Distance) and IBE (Isolation by Environment) analyses, as well as ABBA-BABA introgression inference.


Based on our results, we hypothesize that the evolutionary history of the Royal Irises followed a “stepping-stone” model, where species originated in the north and dispersed down south, followed by an ecological specialization that led to locally adapted populations. Our phylogeny and introgression results suggest that there were two main divergence lines: 1) Iris atropurpurea à I. mariae and 2) I. haynei sensu lato à I. atrofusca. In the south desert region, Iris petrana was the result of a hybridization event between I. atrofusca and I. mariae.
This study is currently under revision in Molecular Ecology.
Genetic basis of flower size in irises (work in progress)
We are analyzing differential expressed genes between two populations of Iris atropurpurea (Large flowers: NET, Small flowers: KUR).
We DE Novo assembled a transcriptome from a total 25 samples (6 genotypes per population). Based on the flower size, we classified three groups: SMALL, MEDIUM, LARGE. We performed DE analyses between those groups. Here are some preliminary results:


To be done: BLASTX, GO terms for gene function, KEGG analyses and SNPs.
EXTRAS AT TAU
During my time at Tel Aviv University, I was in charge of maintaining the Royal irises collection. We collected some juvenile plants and planted them at the Botanical Garden at TAU.



Color polymorphism in Anemone coronaria








As a fun thing, I am helping with some observations of natural populations and cool experiments to answer questions related to pollinators preferences.
Anemone coronaria, better known as poppy, has mainly red flowers but also pink, purple or even white ones are present and are pollinated by bees and beetles. In the lab, my colleagues wonder whether bees prefer a certain color, observations and literature suggest that bees only visit the red flowers; while no preferences are expected for the beetles. I helped a bit on a transplanting experiment to test this hypothesis. I look forward to see what are the results 🙂





Is there a relationship between color polymorphism and seed dispersal ability?
Together with PhD student Tzlil Labin and Dr. Jinlei Zhu, we are studying the dispersal abilities of Anemone coronaria based on Jinlei´s dispersal model and evaluating whether populations with different color morphs have some differences in their seed dispersal.
This work is in progress, hopefully some exciting results will come soon 🙂
Postdoctoral project at Botany Institute at Czech Academy of Science
(Population biology group)
In-situ and ex-situ conservation of Minuartia smejkalii








During my time as a postdoc at the Botany Institute in Pruhonice (close Prague), I was involved in the conservation program of the endemic Minuartia smejkalii.
Potential risk of interspecific hybridization in ex situ collections
In this project we evaluated the consequences of ex situ hybridization between Minuartia smejkalii and a sister species Minuartia caespitosa. We found that hybrids can grow and reproduce faster than the parental species and if they are introduced by mistake in nature they could outcompete the focus endemic Minuartia smejkalii.
These two species are very similar morphologically, and we can only distinguish them observing the structures of the seeds.
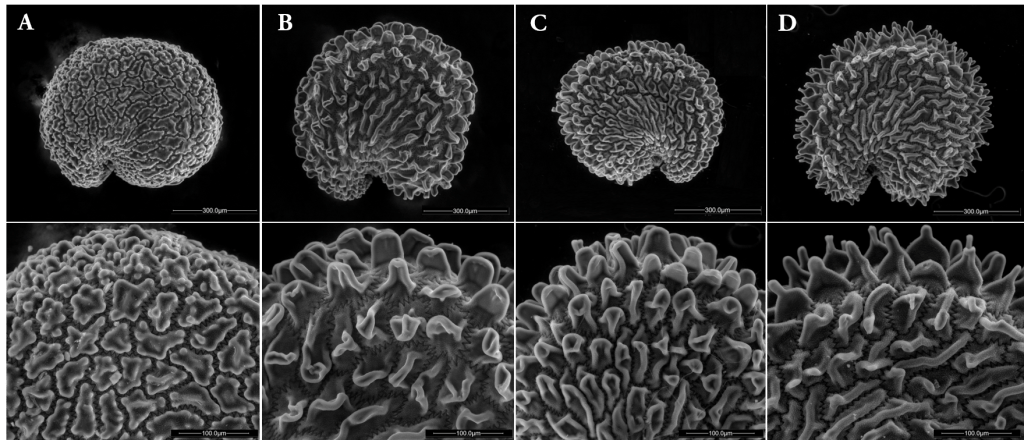
Our results showed that M. smejkalii and M. caespitosa can hybridize and produce viable seeds. Hybrids had a higher germination rate, higher number of flowers in open sites and serpentine soils, and higher dispersal potential.
Our results provide evidence that hybrid individuals from ex situ collections might outcompete the endemic M. smejkalii in the wild, if they are introduced by mistake. So to achieve a successful reintroduction of M. smejkalii into the wild, hybridization in ex situ facilities should be avoided.
A pre-recorded talk presenting the main results about the consequences of ex-situ hybridization can be found here https://vimeo.com/493766996.
Interactive effects of light, water, soil type and competition on Minuartia smejkalii vary over time
We also studied the effect of competition and abiotic conditions and how they change over time. We analyzed the effects of soil type, water, light and competition on the performance of Minuartia smejkalii (plant size, number of flowers) during 4 years (2017-2020).
In a full factorial experiment:
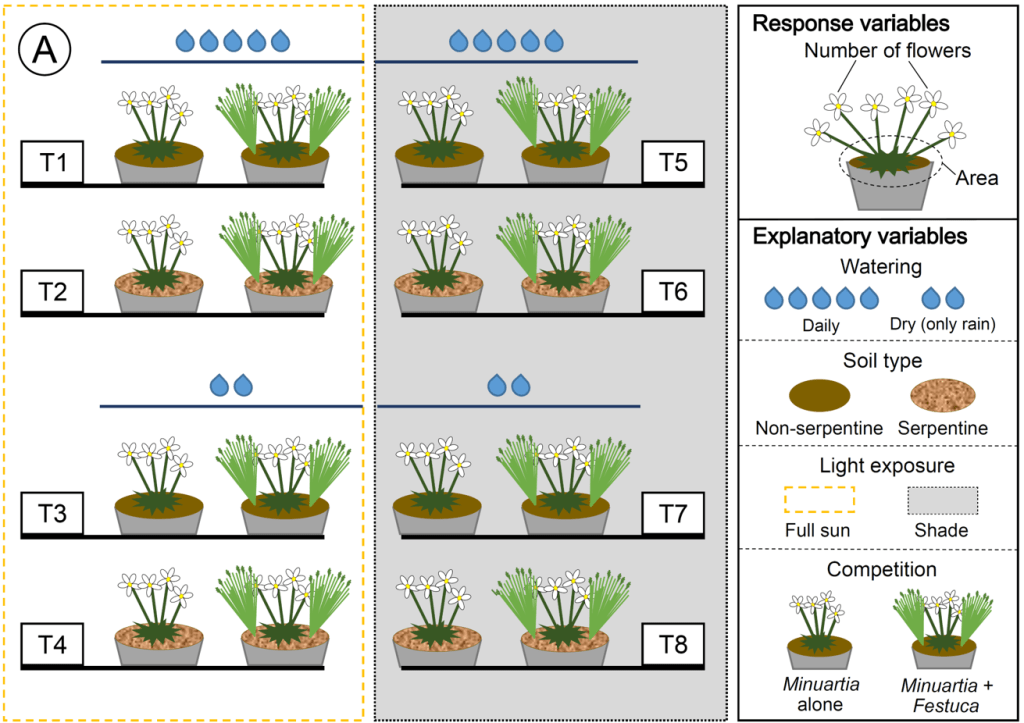
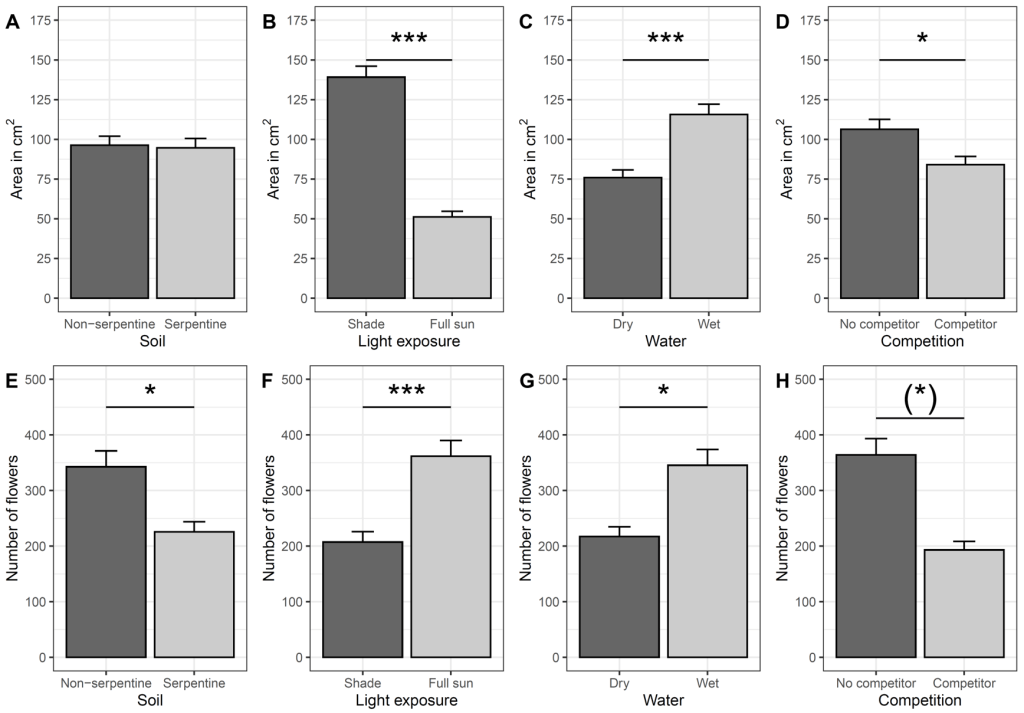
We found interactive complex effects and performance of M. smejkalii change over time and depend on soil type, water, light and competition. Competitive interactions mainly depend on light and soil type, with opposite tendencies depending on plant area vs number of flowers suggesting that abiotic conditions modify vegetative and reproductive investment.
To know more about this exciting project visit Zuzana Münzbergová lab at the Institute of Botany in Pruhonice, Czech Republic https://www.ibot.cas.cz/oddeleni-populacni-ekologie/?team=munzbergova-zuzana-2&lang=en and Life for Minuartia http://www.sandwort.eu/
Photos credit: Hanka Panková and Karolína Hrusková. Electronic miscroscope seeds: Jiri Machac.
From genes to communities:
Assessing plant diversity and connectivity in kettle holes as meta-ecosystems in agricultural landscapes
PhD at Potsdam University
Supervisors: Prof. Jasmin Joshi and Prof. Ralph Tiedemann




My PhD was part of the BioMove Graduate School. The main goal of BioMove is to integrate Biodiversity with Movement. More information about BioMove here https://www.biomove.org/.
In my PhD project I focused on plants and how they disperse (seed and pollen) among kettle holes and their diversity patterns as a result of this movement.
The complete thesis can be find here https://d-nb.info/1218405252/34.
Seed dispersal of two wetland species between kettle holes: a model approach
As an extra aside project after finishing my PhD, in collaboration with Dr. Jinlei Zhu we studied the dispersal seed distances of Typha latifolia and Phragmites australis between kettle holes under different scenarios of land-use management. Dr. Zhu has developed a very sophisticated model with different parameters including properties of the plant (release seed height, terminal velocity of the seeds) and also the surrounding vegetation.

High matrix vegetation decreases mean seed dispersal distance but increases long wind dispersal probability connecting local plant
populations in agricultural landscapes
High matrix vegetation decreases mean seed dispersal distance but
increases long wind dispersal probability connecting local plant
populations in agricultural landscapes
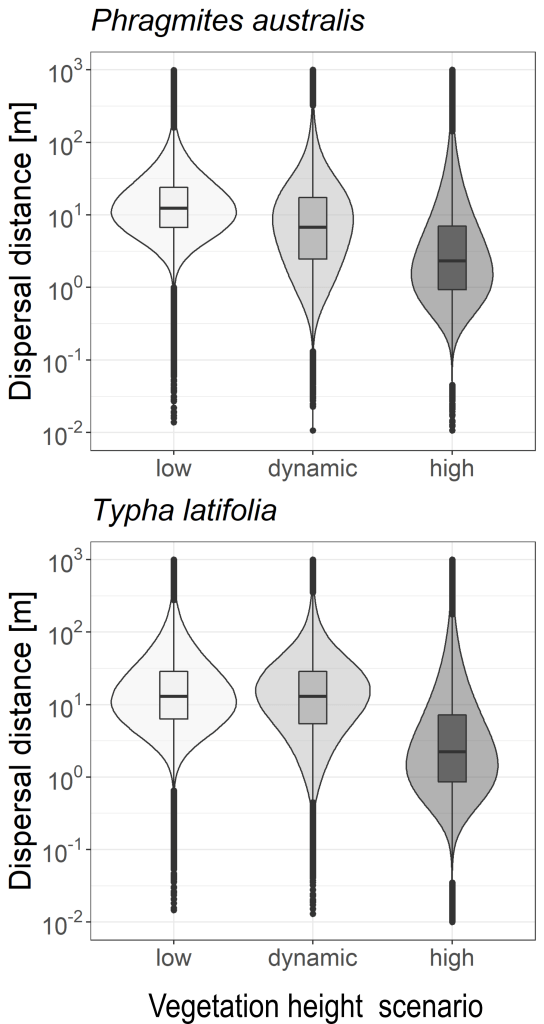


We found that mean seed dispersal distance decreases with the height of surrounding matrix vegetation, but the probability of long-distance dispersal (LDD) increases with vegetation height. This indicates an important vegetation-based trade-off between mean dispersal distance and LDD, which has an impact on connectivity.
The observed positive effect of vegetation height on the probability of LDD and its impact on connectivity provide novel insights into landscape level (meta-) community dynamics — a change in matrix vegetation height by land-use or environmental changes could strongly affect the spread and connectivity of wind-dispersed plants.
More about Dr. Jinlei Zhu work can be found here https://jinleizhu.com/.
Bee diversity in kettle holes

This was the third part of my PhD project. In this project I focused on the bee diversity found in the kettle holes. The idea behind was that these ponds might be important natural habitats for resources (nectar, pollen), where bees can forage and contribute to the pollination in a landscape level.
Habitat quality and connectivity in kettle holes enhance bee diversity in agricultural landscapes
We identified 77 bee species, representing ca. 20% of the total diversity of the Brandenburg region. We found that not only area of kettle holes (rich in floral resources) is important to enhance bee diversity, but also the level of connectivity matters. In addition, habitat quality (i.e. higher flower availability) within the kettle holes positively influenced bee diversity. These results suggested that flowering plants occurring in the kettle holes are an important foraging resource for bees.
Our study identified that a combination of large, high-quality kettle holes providing a high cover of insect pollinated flowers within a high density network of kettle holes, enhances bee diversity in intensively managed agricultural landscapes.
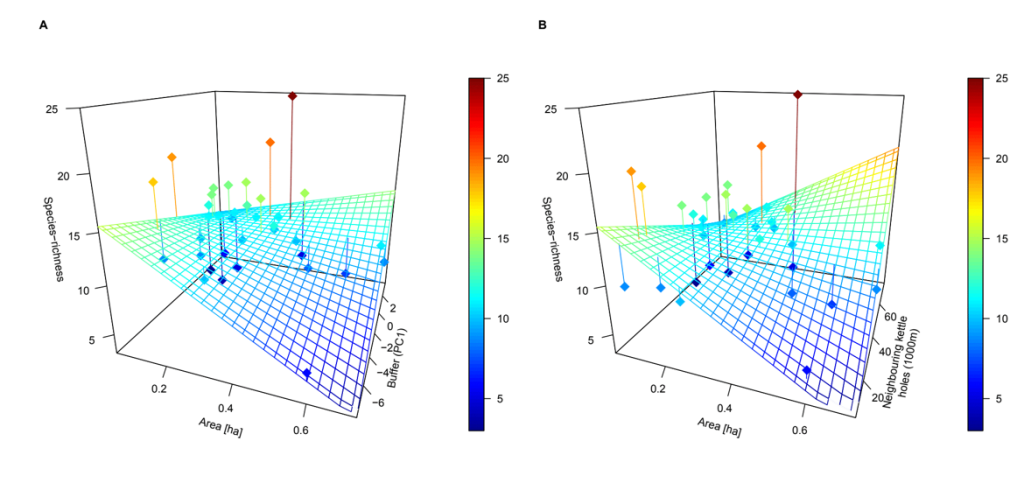
Functional connectivity between kettle holes

This was the second part of my PhD project part of BioMove graduate school at University of Potsdam.
Genetic Diversity and Connectivity in Plant Species Differing in Clonality and Dispersal Mechanisms in Wetland Island Habitats
In this project, we evaluated the degree of connectivity of four wetland species: Lycopus europaeus, Oenanthe aquatica, Typha latifolia and Phragmites australis. They have different movement and reproduction strategies, the first two are mainly pollinated by bees and seeds are dispersed by water, while the last two pollen and seeds are transported by wind. They also differ in the degree of clonality, Typha and Phragmites reproduce almost only clonally, Lycopus less common and Oenanthe almost never.
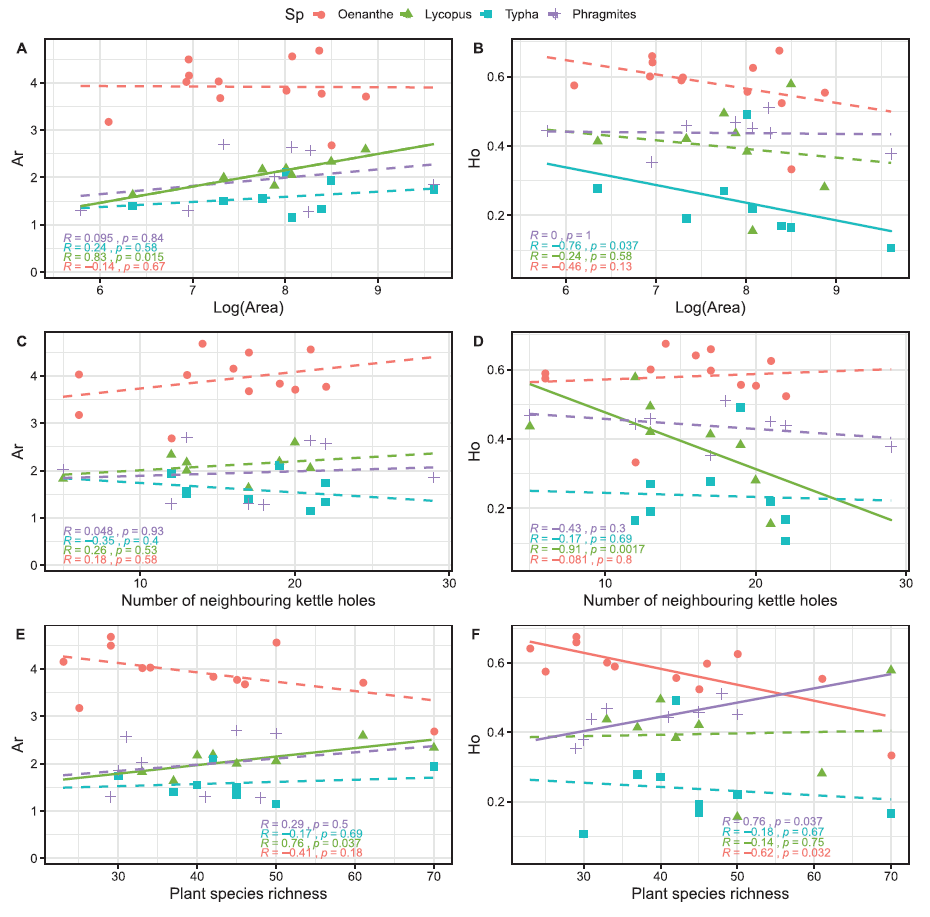
Our results showed that genetic parameters reflect the reproduction strategies with the highest genetic diversity being observed in the non-clonal, outcrossing Oenanthe aquatica compared to the clonal
Lycopus europaeus, Typha latifolia, and Phragmites australis. Even though the non-clonal, outcrossing species showed very high genetic diversity overall, a negative trend with increasing plant species richness suggests locally smaller effective population sizes with increasing interspecific competition. In contrast, genetic diversity in the 2 clonal species Lycopus and Phragmites showed a positive trend with increasing kettle-hole area and with plant-species richness indicating pronounced competitive abilities, probably through rapid clonal expansion.
Plant diversity and dispersal traits in two types of kettle holes: permanent and ephemeral

This was the first part of my PhD. In this study we compared plant diversity occurring in two types of kettle holes: permanent and ephemeral, and also their dispersal traits.
Environmental filtering predicts plant‐community trait distribution and diversity: Kettle holes as models of metacommunity systems
We found that large permanent kettle holes harbor more number of species but the plant composition differed between types of kettle holes. Interestingly, the degree of isolation (number of other surrounding kettle holes) was only important in the ephemeral ones. These results suggest that ephemeral ponds are more dynamic and depend on other surrounding ponds to get plant species. Also in these ephemeral ponds, dispersal traits of the plant communities were more adapted to be dispersed by humans (probably by farmers or agricultural tractors).
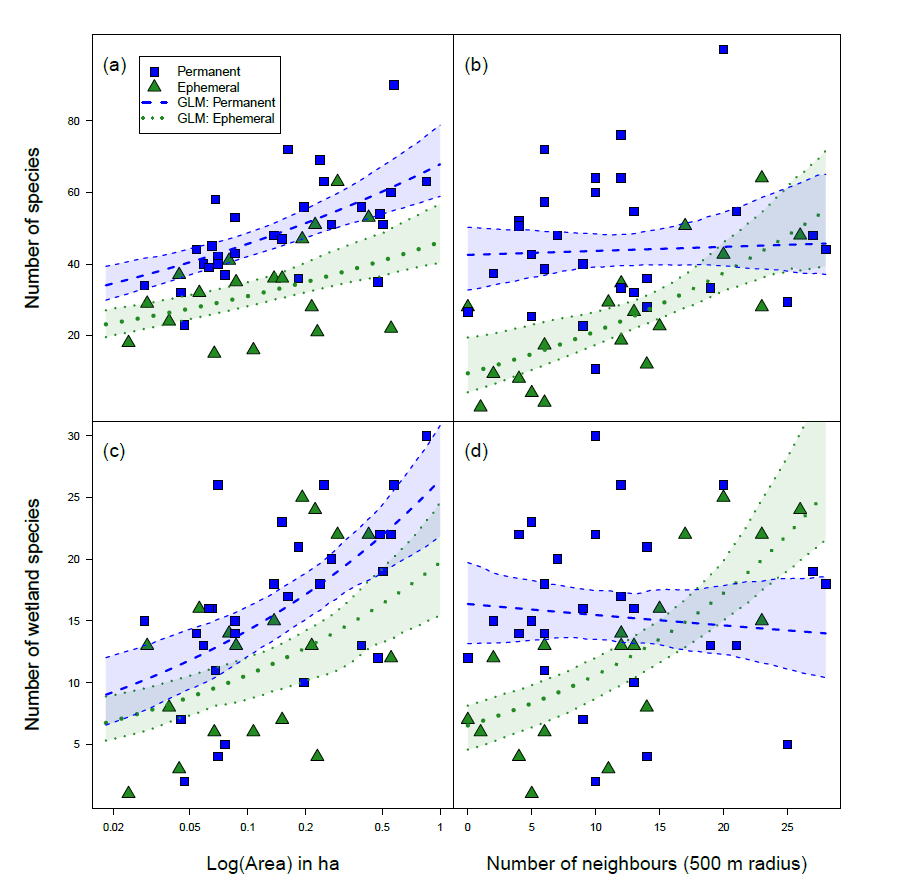
Studies like this show that all different types of ponds are important to get connectivity and movement of species in a landscape level. We recommend to pay attention to the ephemeral types in particular. Ephemeral ponds are threatened since they can dry out some years and farmers would just plough through. They harbor a unique diversity of plants and therefore must be conserved.

Master degree at Bonn University
Flower biology of Impatiens
Analysis of pollination syndromes and breeding systems in Impatiens (Balsaminaceae)
Supervisor: Dr. Stefan Abrahamczyk
My master thesis was divided in two parts: Pollination syndromes and Breeding systems in Impatiens. Impatiens is a genus of plants with more than 1000 species distributed around the world. The genus Impatiens has a high flower diversity adapted to different pollinators. In the picture we can see typical pollination syndromes:

A. Impatiens noli-tangere B. Impatiens niamniamensis C. Impatiens platypetala D. Impatiens sodenii E. Impatiens campanulata F. Impatiens mandrakae. Scale bar 0.5 cm. Photos Sissi Lozada (SL) and M. Ackermann (MA).
A) Bee pollination: Flower with a funnel shape, a landing flat platform for the bees, colored and wide opening to guide the insect towards the thin tube-like structure called spur, where the nectar is located.
B) Bird pollination: Flowers have usually vivid colors like red, are very stable with thick stems to support strong movements by the flying birds. Spur is very wide (only narrow at the tip) containing a lot of nectar with high diluted sugars.
C) Butterfly pollination: Flowers with a broad colored frontal display and a long filiform spur. Butterflies use a long proboscis to suck the nectar located in the tip of the spur.
D) Moth pollination: Flowers similar to butterfly pollination in shape but usually are not very bright, with longer spurs and aromatic to attract moths at night.
E) Generalist pollination: Flower pollinated for more than one group bees, butterflies and howkmoths.
F) Fly pollination: Small colorless flowers with no spur and no nectar.
Recently in the Trimorphopetalum clade which are endemic species from Madagascar, we report the first case of deceiving fly pollination. In the open-lipped flowers of Impatiens rivularis endemic from humid forests in Madagascar produce a fungi scent that attract small flies. During their life cycle, these flies usually lay eggs in fungi, the flower deceive them with the fungi scent and when the flies are looking for a place to lay eggs, they probably get pollen on their bodies and transport to another individual. Fascinating!

Impatiens rivularis